Category: life extension – Page 648
Human Skeletal Muscle Aging and Mutagenesis
An article I wrote:
Study based upon human skeletal muscle aging, mutagenesis, and the role of #satellite cells
“A more comprehensive understanding of the interplay of stem cell–intrinsic and extrinsic factors will set the stage for improving cell therapies capable of restoring tissue homeostasis and enhancing muscle repair in the aged.”
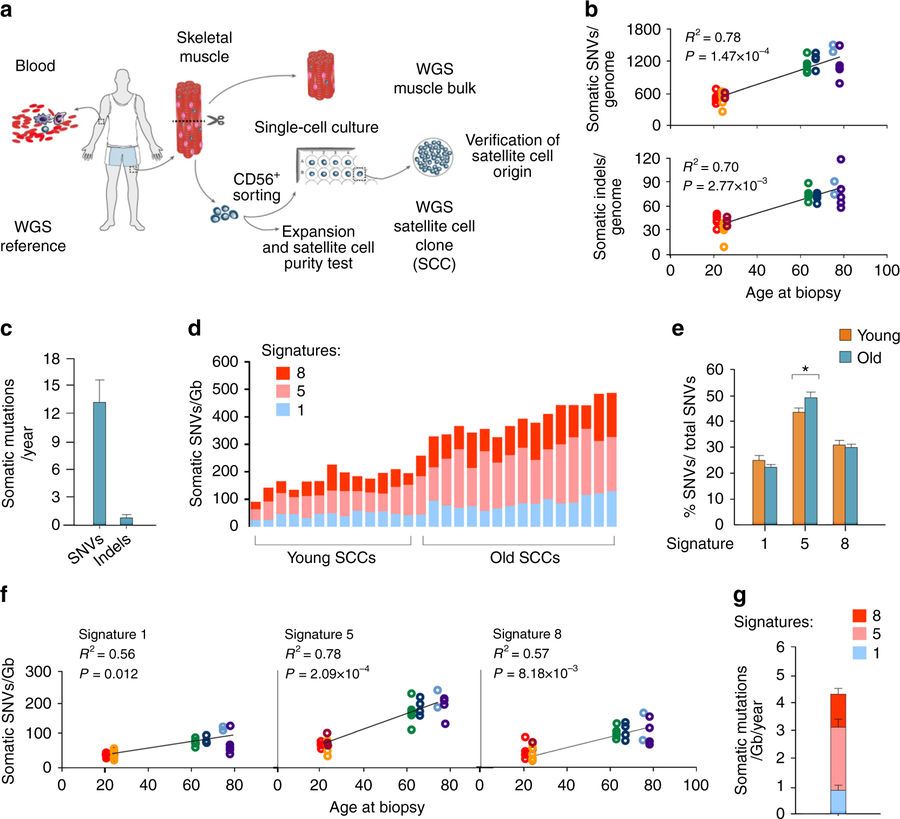
Human aging has multiple effects on the human body. One of the effects of human aging is the reduction in skeletal muscle (SkM) function and a reduction in the number and activity of satellite cells (SCs), the resident stem cells. The whole genome of single SC clones of the leg muscle vastus lateralis from healthy individuals of different ages (21–78 years) was analyzed, to study the specific connection between SC aging and muscle impairment. In healthy adult muscle rapid increase of SCs is consistent with the accumulation rate of 13 somatic mutations per genome per year. Mutations typically do not happen in SkM-expressed genes because they are protected. However, as mutations in exons and promoters increase, genes involved in SC activity and muscle function are targeted which results in aging. Exons are coding sections of an RNA transcript, or the DNA encoding it, that are translated into protein. Proteins are the synthesis of molecules.

Inevitable muscle wasting of old age could be stopped, scientists believe
T he inevitable muscle wasting of old age could be stopped, scientists believe, after discovering why people become frail as they grow older.
Most people become weaker in their later years, as their leg muscles get smaller and less able to bear weight, which often leads to disability and falls.
But until now, nobody has known why the process happens or if it can be reversed.

What if billionaires could live forever?
Several billionaires, most of them Californians, have been funding firms involved in developing life-extension technologies. What if they succeed? What if billionaires alive today live indefinitely and get ever richer?
February saw the announcement in Silicon Valley by X Prize founder, serial entrepreneur, and all-round gee-whiz future-technology promoter Peter Diamandis that he had cofounded a new company called Celularity.
He did so together with Dr. Bob Hariri, a renowned biomedical entrepreneur known for innovations in harvesting placental stem cells. Hariri had previously founded Celgene Cellular Therapeutics.
Placenta makes 100-years old the new 60 — Fox Business Mornings With Maria
Celularity, a leading U.S. biotechnology company, is seizing on an opportunity to use cells to target diseases. The CEO, Dr. Robert Hariri, says placenta can be used to augment longevity and immunity. https://www.celularity.com
Celularity’s Massively Transformative Purpose (MTP) is to harness the power of the living cell to augment biology, immunity and longevity.
/redirect?q=http%3A%2F%2Fwww.foxbusiness.com%2Ffeatures%2F2018%2F02%2F15%2Fbiotech-breakthrough-placenta-makes-100-years-old-new-60-research-finds.html&v=jkQV3gUua_8&event=video_description&redir_token=QKlLB38bGZR8iE-br1upaZX8KUh8MTUyMDg4MzgwNEAxNTIwNzk3NDA0
Enzymes and Cognitive Decline
Enzymes play an important role in cognitive function. Enzymes are biological catalysts. They’re responsible for accelerating chemical reactions.
What role do enzymes play in #aging and cognitive function?
According to new research in laboratory mice by UC San Francisco scientists have discovered that loss of an #enzyme that modifies gene activity to promote brain regeneration may be partly responsible for age-related cognitive decline. When age related cognitive decline starts is still debatable, however the effects of age related cognitive decline are well known.
Suicide Gene Therapy Works to Kill Cancer Cells
Some cancer cells express some of the same genes that senescent cells do, so it makes sense that drugs that destroy senescent cells may also destroy cancer cells. This was what the researchers in this new study set out to test.
However, in this experiment, the researchers discovered that the chosen senolytic drugs were not effective at destroying cancer cells with senescence-associated gene expression. While cancer cells and senescent cells do share some common properties, they are also quite different at an epigenetic level.
The researchers did, however, demonstrate that a so-called “suicide gene therapy” that causes both senescent cells and cancer cells to kill themselves worked by targeting senescence-associated p16Ink4a. This approach is similar to that of SENS spin-off company Oisin Biotechnologies, which is using a suicide gene therapy to eliminate senescent cells.
Can humans live to be 1,000 years old?
And what the media and scientists think of it.
Aubrey de Grey, Ph.D., Vice President of New Technology Discovery at AgeX Therapeutics, discusses the “Methuselarity” — the point at which technology enables humans to live to more than 1,000 years of age. Dr. de Grey believes this could happen within the coming decades and posits that some people born today may live to be 1,000 years of age. He further states that people who are 30 years old today have a 50/50 chance to live to be 1,000 years old. Dr. de Grey bases his assumptions on the research into aging that companies like Agex Therapeutics are pursuing. This video is the third in a series from AgeX about the future of aging and its impact on humanity. For more information on the company, please visit www.agexinc.com.

Bioquark Inc. — Conferences on Death / Death Reversal — Ira Pastor
Two pivotal conferences on the topic of “death” coming up!!
First at the INSERM Liliane Bettencourt School on March 16–18 will be “Death: From Cells to Societies — Aging, Dying, and Beyond” -
Then, April 11–13 at Harvard Medical School, will be “Defining Death: Organ transplantation and the 50-year legacy of the Harvard report on brain death”
http://bioethics.hms.harvard.edu/annual-bioethics-conference-2018
An important inflection point for all!!